
©
Leandro Castelluccio
Introducción
El sistema de la visión ha sido ampliamente estudiado dentro del campo de la Neurociencia y las Ciencias Cognitivas. Es un área fascinante y a la vez que compleja, desde donde surgen multitud de insights acerca del funcionamiento cognitivo y cerebral en general, brindando nuevas posibilidades de entendimiento del sistema nervioso, así como de nuevas aplicaciones en el campo de la Neurociencia Computacional. En este ensayo daremos una breve reseña del sistema de la visión, las ilusiones ópticas y qué implican las mismas para entender lo que el cerebro hace cuando procesa los estímulos sensoriales (¿predice activamente o se activa pasivamente?).
El sistema de la visión
Uno de los elementos básicos del funcionamiento cerebral radica en la sinapsis. Este es el punto donde las señales eléctricas de una neurona generan efectos eléctricos en otra. La iniciación de un potencial de acción, por ejemplo, aquello que permite transmitir impulsos nerviosos, y que es la base del funcionamiento cerebral a nivel de la neurona, requiere de la apertura de canales iónicos de sodio en el montículo del axón de la neurona. A este nivel, los fenómenos dependen de propiedades físicas y químicas y de las características de las estructuras biológicas en juego. Para desatar un potencial de acción, un determinado umbral de disparo debe ser superado gracias a la acción de potenciales graduados que se suman temporal y espacialmente, provistos por las conexiones con otras neuronas. Para que se geste actividad en estas últimas deben ocurrir los mismos fenómenos (Kandel, Schwartz,& Jessel, 2001).

El proceso de la visión es un buen ejemplo para entender lo que significa la percepción a nivel de los órganos de los sentidos y el procesamiento cerebral. A nivel de la retina, en nuestros ojos, se encuentran dos tipos de fotorreceptores: los conos y bastones. Los fotorreceptores son células sensibles a la luz que transducen la misma en una señal eléctrica. Los conos son fotorreceptores sensibles en condiciones fotópicas (luz diurna). Hay 3 tipos: S (sensible a la longitud de onda corta), M (sensible a la longitud de onda media) y L (sensible a la longitud de onda larga). Los bastones, por otro lado, son fotorreceptores sensibles en condiciones escotópicas (oscuras). Los bastones, por ejemplo, poseen un pigmento visual denominado rodopsina, el cual presenta dos partes, una porción proteica ubicada en la membrana de los bastones, y una retinal, derivada de la vitamina A. Esta última parte posee dos configuraciones isométricas distintas: 11-cis y hola-trans. La luz, a través de un proceso físico, lo que hace es cambiar la configuración de esta parte de la rodopsina de 11-cis a holo-trans, y aquí se inicia una cascada de acontecimientos que terminan en la experiencia de visión que tenemos (Kandel, Schwartz, & Jessel, 2001).
Otro concepto clave es el de la fóvea, un área en la retina central en el centro de la mirada donde los fotorreceptores están más compactos y la agudeza visual es más alta. La retina representa una superficie neural sobre la cual el ojo proyecta una imagen. Contiene varias capas de procesamiento que terminan en las células ganglionares. Antes de eso, también tenemos las células amacrinas, que son células laterales de procesamiento en la retina, para las cuales hay más de 40 tipos anatómicamente distintos. Por otro lado, las células horizontales son células de procesamiento lateral que subyacen a la organización central-envolvente de los campos receptores bipolares y de las células ganglionares. Un concepto clave aquí es de los campos receptivos, que tienen una estructura central-envolvente. El campo receptivo de una neurona visual es el área de la retina que, cuando es estimulada por la luz, causa una respuesta en la neurona. Luego también tenemos las célula bipolar, que es una célula de procesamiento de la retina que transporta una señal eléctrica desde los conos hasta las células ganglionares. Finalmente, las células ganglionares son neuronas de salida de la retina, que se conectan al NGL (núcleo geniculado lateral, que transmite información visual desde la retina a la corteza) a través del nervio óptico. Sus campos receptivos tienen una estructura central-envolvente. Hay más de 30 tipos diferentes, cada uno sintonizado a información visual diferente. Las células enanas, por ejemplo, son células ganglionares especializadas para alta agudeza y color “rojo-verde”. Las células del parasol, por otro lado, son células especializadas para altas frecuencias temporales y bajas frecuencias espaciales. Otro ejemplo son las células pequeñas bistratificadas, células ganglionares de la retina especializadas para el color violeta-cal (Kandel, Schwartz, & Jessel, 2001).

Siguiendo a estos autores, dos conceptos clave son el campo receptivo de la neurona visual, que es el área de la retina, que, cuando es estimulada por la luz, causará una respuesta en la neurona, y la noción de células de centro y fuera del centro. Las células de centro se refieren a aquellas en las que la luz que cae en el centro del campo receptivo es excitadora y la luz que lo rodea es inhibitoria. Las células de fuera de centro se refieren a aquellas en las que la luz que cae en el centro del campo receptivo es inhibitoria y la luz que cae en el entorno es excitadora.

Y luego nos movemos a un nivel superior de procesamiento A nivel cortical en el cerebro, el proceso de la visón se organiza en dos vías desde los lóbulos occipitales: la dorsal, dirigida hacia la corteza parietal posterior, y la ventral dirigida hacia la corteza temporal inferior. La vía dorsal implicaría el procesamiento de elementos como la profundidad, el movimiento y la localización en el espacio de los objetos captados. La vía ventral implicaría el procesamiento de elementos como la forma o el color. Cada núcleo de procesamiento en el cerebro respondería a aspectos particulares de la imagen captada, donde un grupo neuronal específico se activarían en función de determinado aspecto de la imagen, como determinado color, por ejemplo, pero no para otro aspecto, como las formas o contornos de la imagen. El procesamiento visual representa una codificación dispersa, donde la presencia de un estímulo está codificada por la actividad de una red de neuronas dispersamente distribuida. Conceptos clave son los del sistema magnocelular y el sistema parvocelular. Se cree que el sistema magnocelular, que se origina a partir de las células de parasol sintonizadas con frecuencias temporales altas y bajas espaciales, da lugar al “sistema donde”. El sistema parvocelular, por otro lado, refiere a un flujo visual que se origina en las células enanas que se cree que dan origen a la ruta “qué”. La ruta “donde” representa los flujos de información desde la corteza visual primaria (un área que contiene células que prefieren líneas con orientaciones diferentes, diferentes frecuencias espaciales y regiones de “burbujas” sensibles al color) a través de otras áreas visuales, incluida la MT (área visual especializada para el movimiento) al lóbulo parietal y está relacionada con la ubicación y el movimiento (la vía dorsal antes mencionada). La ruta “qué” representa los flujos de información desde la corteza visual primaria a través de otras áreas visuales, incluida la V4 (especializada en color) hasta el lóbulo temporal, y está relacionada con la forma y el color (la vía ventral) (Kandel, Schwartz, & Jessel, 2001).
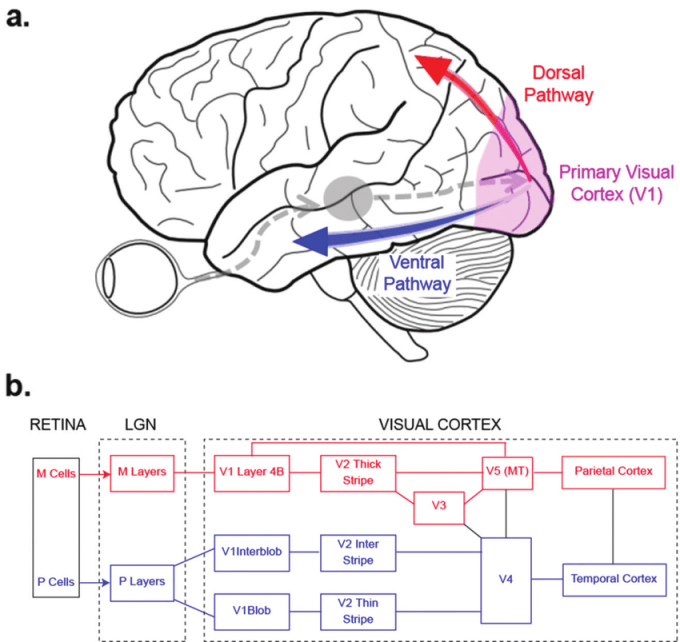
La figura de arriba muestra un ejemplo esquemático de la organización del sistema visual. Sin embargo el mismo es mucho más complejo de lo que clásicamente se ha considerado, con interconexiones entre varios núcleos de procesamiento, tal como ejemplifica el esquema de Felleman y Van (1991):

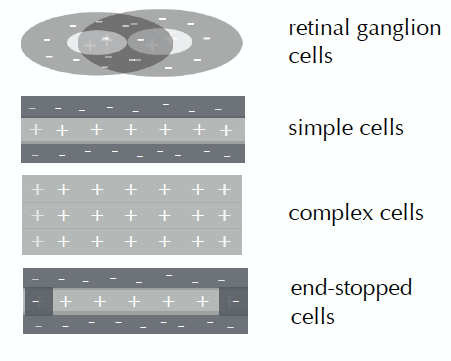
Siguiendo a Kandel, Schwartz y Jessel (2001), dentro de la corteza visual, las células simples representan células de la V1 (corteza visual primaria) con campos receptivos que prefieren bordes orientados. Las células complejas son células encontradas en V1 y V2 (corteza visual secundaria) que son similares a las células simples pero con un grado de invariancia espacial. El borde podría caer en cualquier parte del campo receptivo o podría estar moviéndose y la célula todavía respondería. Células “detenidas-final” (end‐stopped cells en inglés) son neuronas en V1 y V2 que responden a líneas orientadas de una longitud particular. Las células simples responden a los bordes. Sus campos receptivos se pueden mapear en zonas fijas excitatorias e inhibitorias. Las células complejas también responden a bordes orientados, pero tienen un grado de invariancia espacial: responden a un patrón de luz de una orientación dada, independientemente de dónde se encuentre exactamente en el campo receptivo. Por lo tanto, responde a un estímulo en movimiento y puede ser direccionalmente selectivo para el movimiento. Las células simples y complejas pueden tener campos receptivos que se detienen al final, lo que significa que la respuesta se reduce para bordes más largos que una cierta longitud. Los campos receptivos más complejos se crean al cablear células que tienen campos receptivos más simples:
Cuando ocurren lesiones a nivel de la corteza visual surgen distintos síndromes, tales como la Acromatopsia o la Ceguera Cortical (Blindsight). La Acromatopsia o la Ceguera Cortical al color se producen cuando el mundo se percibe de forma completamente acromática pero los conos funcionan normalmente. Es causada por una lesión cerebral localizada que a menudo incluye V4 y/o V8. El Blidsight, por otro lado, se refiere a la ceguera cortical causada por una lesión en la corteza visual primaria. Todavía es posible cierta percepción visual, particularmente del movimiento por una vía subcortical, pero los pacientes con visión ciega pueden tener visión sin conciencia.
Las ilusiones ópticas: ¿por qué suceden?
Un factor clave para entender las ilusiones ópticas es la noción de constancias. Por ejemplo, la constancia de tamaño, refiere a percibir la propiedad de tamaño de un objeto como constante independientemente de la distancia o ubicación del mismo. Por otro lado, la constancia del color es la percepción constante del color de un objeto a pesar de los cambios en el color de la iluminación del objeto y, por lo tanto, en el color de la luz que refleja el mismo. Estos son solo una serie de constancias, que también incluyen la forma y la rugosidad, por ejemplo. Estas constancias nos permiten objetivizar nuestra realidad, es decir, nos permite ver las cualidades de los objetos respecto de los cambios y las condiciones, lo que forma una imagen clara del mundo exterior y los tipos de entidades que conforman la realidad (Gilchrist, 2010). Como hemos visto anteriormente, el sistema visual se puede dividir en tres niveles de procesamiento de información: visión de nivel bajo, que se centra en los mecanismos fisiológicos de la retina y las señales neurales resultantes; visión de nivel alto, que es cognitiva e implicaría conocimiento previo; y un nivel medio que a veces se asocia con la psicología de la Gestalt y un énfasis en las estructuras organizativas de la percepción (Adelson, 2000). Tanto el procesamiento de nivel bajo como el nivel alto son factores que están asociados a ilusiones ópticas comunes. El procesamiento de nivel bajo, como en la retina y los procesos corticales tempranos, proporcionan un mecanismo subyacente para la constancia de la luminosidad, por ejemplo, donde nuestro sistema visual es menos sensible a los cambios graduales en la luminancia que a los contrastes locales repentinos, y las regiones uniformes se “completan”. En ciertos casos perceptivos, este factor genera una ilusión óptica. Sin embargo, algunas explicaciones de bajo nivel no han podido dar cuenta de las ilusiones y se han formulado explicaciones de nivel alto. En el nivel alto, la percepción podría considerase como el resultado de inferencias inconscientes. La constancia de luminosidad, por ejemplo, se lograría mediante el sistema visual inferiendo y luego descontando el iluminante. Las ilusiones en estos casos son el resultado de una falla en nuestras inferencias inductivas de lo que hay en el mundo. Por lo tanto, al utilizar premisas basadas en evidencia sensorial, condiciones de iluminación percibidas y experiencias anteriores, nuestro sistema visual llega a una conclusión falsa. Sin embargo, también debemos señalar que parece que no existe prueba empírica para la existencia de inferencias inconscientes, y que las teorías modernas de la percepción de la luminosidad incorporan mecanismos de alto y bajo nivel para explicar nuestra experiencia (Kindom, 2011).
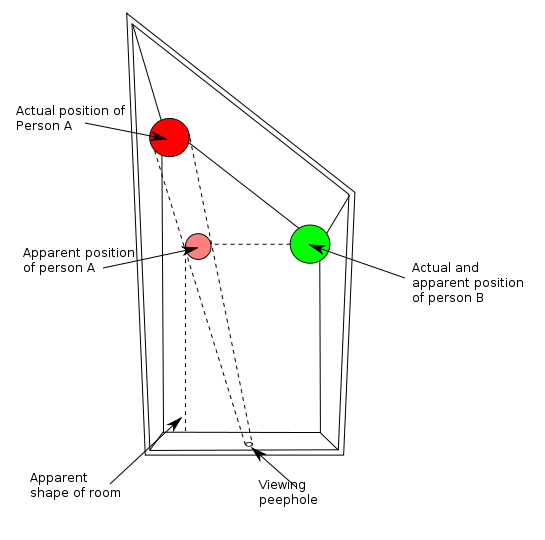
Una ilusión óptica clásica es la de “la habitación de Ames”, una ilusión en la que dos figuras parecen de diferentes tamaños:

Tal como explica este video, una persona está más alejada del observador que la otra, pero las señales en la habitación están arregladas para parecer que estan a la misma distancia. Por lo tanto, la constancia de tamaño no se aplica al estímulo más distante para adecuar su tamaño percibido. En este ejemplo se engaña al observador modificando las características normales que tendría una habitación y jugando con el ángulo de la cámara que toma la imagen de dicha habitación:
Otro ejemplo es la ilusión de la cuadrícula, una cuadrícula cuadrada donde se perciben manchas ilusorias en intersecciones brillantes:

Una explicación de este fenómeno toma en cuenta el nivel bajo de procesamiento visual, haciendo referencia al campo receptivo visual, donde como mencionábamos, en el centro del campo, los fotorreceptores individuales excitan la célula ganglionar cuando detectan un aumento de luminancia mientras que los fotorreceptores en el área circundante inhiben la célula ganglionar. Por lo tanto, dado que un punto en una intersección está rodeado de más intensidad que un punto en el medio de una línea, la intersección aparece más oscura debido a la mayor inhibición:

Sin embargo, si realizamos un análisis de Fourier, donde cualquier función se puede expresar como la suma de una colección de sinusoides y transformamos la cuadrícula, la ilusión desaparece, aunque sigue existiendo el mismo procesamiento visual de nivel bajo, lo que indica que un probable proceso visual de nivel superior es requerido para explicar la ilusión:

Por otro lado, tenemos la ilusión de sombra a cuadros, donde se percibe una diferencia entre la claridad de dos superficies idénticas. La ilusión parece ser producto de la constancia de la luminosidad:

Como indica la siguiente imagen, en realidad tanto el cuadrado A como el B tienen el mismo tono de gris:

La constancia de luminosidad se logra infiriendo y luego descontando el iluminante. Si bien hay una sombra representada a través del tablero de ajedrez, en realidad no hay una sombra allí, ya que en la ilusión solo hay una imagen de una sombra, lo cual causaría la ilusión. Dicha ilusión ejemplificaría el resultado de una falla en nuestras inferencias inductivas.
Por otro lado, una ilusión interesante descripta por Tangen, Murphy y Thompson (2011) muestra un nuevo efecto de distorsión de la cara, que resulta de la presentación rápida de caras alineadas a los ojos. Estos autores reconocen la experiencia como “cada cara se convierte en una caricatura de sí misma, algunas caras parecen muy deformes, incluso grotescas” (la traducción es mía del inglés de: Tangen, Murphy y Thompson, 2011, página 628). Aparentemente, el grado de distorsión es mayor para las caras que se desvían de las demás en el conjunto en una dimensión particular: si una persona tiene una frente grande, parece particularmente grande, si alguien tiene una nariz delgada, entonces se ve notablemente delgada o si una persona tiene una mandíbula grande, parece particularmente grande, casi como un ogro. Observe el siguiente video para ver un ejemplo de esta ilusión.
Según estos autores, la codificación relativa parece impulsar el efecto. Esto significa forzar al observador a codificar cada cara a la luz de las otras, y al alinear las caras con los ojos, resulta mucho más fácil comparar su forma y la ubicación relativa de sus características, haciendo que las diferencias entre ellas sean más evidentes. La velocidad de presentación rápida y constante también puede fomentar esta codificación relativa, evidente por el hecho de que el efecto disminuye si las caras no están alineadas a los ojos o se presentan demasiado lento o demasiado rápido. Además, si se inserta un breve espacio en la secuencia, el efecto desaparece casi por completo (Tangen, Murphy, & Thompson, 2011)
Una ilusión similar, el “efecto de distorsión de la cara después del efecto” podría indicar que la adaptación es clave para generar el efecto de distorsión de la cara (Webster & MacLin, 1999). En un estudio realizado por estos autores, los observadores igualaron o clasificaron las caras antes o después de ver imágenes distorsionadas de las caras, lo que demuestra que la adaptación anterior influye fuertemente en la percepción de la cara al hacer que la cara original aparezca distorsionada en una dirección opuesta a la adaptación de la distorsión. En su experimento, los efectos secundarios fueron más débiles cuando las caras de adaptación y de prueba tenían diferentes orientaciones, concluyendo que los efectos secundarios dependen de qué imágenes están distorsionadas y no simplemente del tipo de distorsión introducida. Además, según Hills, Holland y Lewis (2010), los niños (de 6 a 12 años de edad) mostraron mayores efectos secundarios que los adolescentes (de 13 a 18 años de edad) y demostraron efectos posteriores de una magnitud similar para las distorsiones asimétricas y simétricas, sin embargo los adolescentes solo mostraron efectos secundarios por distorsiones simétricas. Estos autores proponen que los niños pueden tener una norma facial y respuestas neuronales más flexibles, que permiten una gama más amplia de estados adaptados en comparación con los adolescentes, lo que implica aún más la adaptación como un mecanismo para el efecto generado. Esto implicaría que los sistemas visuales se basan en información aprendida anterior y en la flexibilidad actual para procesar estímulos visuales, lo que podría apoyar la noción de visión como codificación predictiva.
Según Tangen et al. (2011), el efecto de distorsión parece depender de las dimensiones exteriores entre las imágenes del conjunto, y estas dimensiones no parecen estar limitadas a las características o configuraciones faciales, si una fotografía está muy iluminada en comparación con las demás, entonces aparece sobreexpuesta. Pero en general, el mecanismo bajo el cual se produce el efecto no se comprende del todo.
¿Codificación predictiva?
Tal como destacaba en un ensayo previo (“Aplicando los conceptos de Bayes al entendimiento de la conciencia – Algunos errores“) existen muchos casos de ilusiones ópticas visuales que podría darnos la idea de que el aspecto consciente de nuestra experiencia es un proceso de inferencia del cerebro de lo que está afuera de este en el ambiente y no un exacto reflejo de la realidad (aunque nunca en realidad lo sea, hablamos aquí de ciertos errores como ocurre en ciertas ilusiones ópticas). Existe una ilusión que puede ser un ejemplo de esto, la máscara giratoria: link.
Dicha ilusión se basa en el procesamiento del rostro humano. El tema aquí no tiene que ver con el sentido del giro sino con lo que parece ser una imposibilidad de percibir rostros huecos. El cerebro parece estar condicionado a ver rostros normales, lo cual es muy interesante, porque es una ilusión, entre otras, que podría considerarse como argumento de la conciencia como un producto de tipo bayesiano: lo que percibimos conscientemente no son percepciones totalmente objetivas, por decirlo de una manera, lo que la persona ve, sino predicciones del cerebro de lo que debería estar viendo (codificación predictiva). Tal como en la muestra la ilusión de sombra a cuadros antes mencionada, el cerebro asume que hay una sobre y eso condiciona la percepción del cuadrado B. En la ilusión del rostro hueco, cuando el rostro se va dando vuelta y queda expuesta la parte hueca, en ese momento, el cerebro parece no procesar el rostro hueco e invierte la imagen del rostro para verlo normal.
Pero lo interesante de estas ilusiones en relación a la codificación predictiva, es que no se toma en cuenta el carácter visual de las mismas, que puede por sí solo explicar las ilusiones, esto ha sido pasado por alto en relación a la conciencia como un producto de inferencia. ¿Qué quiero decir con esto? Que si tales ilusiones visuales hablan de un carácter predictivo del cerebro, se pasa por alto otros tipos de percepciones que deben confirmar la regla, ¿por qué no hay tales ilusiones diversificadas en modalidades sensoriales como el tacto o el sentido del olfato? ¿Acaso no será más factible que existe una malinterpretación de estas ilusiones en cuanto a que implican una codificación predictiva?
Como mencionaba en el ensayo previo respecto a Bayes, pensemos cuando vamos en bus o tren junto a otro, y estando detenidos el otro comienza a moverse pero parece que es uno el que lo hace, podríamos preguntarnos: ¿está el cerebro condicionado o predispuesto a verse uno mismo moviendo en vez de otras cosas? Lo cierto es que a veces parece una cosa y a veces otra. En tal caso el cerebro no espera que sea uno el que se mueva, de hecho contempla ambas posibilidades, y sin embargo esa ilusión se da y el cerebro no tiene la expectativa de que es uno el que se mueve, no existe ese “a priori”. De tal forma, la ilusión obedecería a cuestiones perceptivas que van más allá de un funcionamiento predictivo del cerebro netamente. Y de nuevo, si fuera una codificación predictiva en términos absolutos para el cerebro, ¿por qué no podemos corregir los errores de predicción en las ilusiones visuales incorporando nueva información de carácter perceptivo? ¿No sería esto más adaptativo en término evolutivos? Pues, si estas ilusiones ópticas fueran resultado de un producto de tipo bayesiano, las ilusiones deberían cambiar luego de que tenemos un nueva información de sus caracter erróneo como en el caso de la ilusión de sombra a cuadroso en la ilusión de la máscara hueca. Pero esto no sucede. Tal como indican Ramachandran yHirstein (1997), los qualia (nuestra experiencia subjetiva consciente) presentan tres leyes. El ser humano exhibe ciertos procesos que son irrevocables, el quale sería uno de ellos. La primera ley indica así que los qualia son irrevocables, esto es, una vez que uno ha identificado una sensación, uno no puede rechazarla, una vez que uno percibe un objeto de color azul, uno no puede verlo de otro color, por más que uno lo desee conscientemente. Por lo que la experiencia tiene un carácter definido que sencillamente no puede cambiar. Esto contradice la posibilidad de que podamos ajustar nuestras percepciones directas de las cosas, ciertos qualia, dada nueva información que ajuste nuestras predicciones o los “a priori”. De modo que por más que quisiéramos ver los dos cuadrados con el mismo color de gris en el ejemplo anterior de la ilusión de la sombra, no podemos hacerlo, aunque hayamos visto que tienen el mismo color. Esto indicaría que la conciencia va más allá de una predicción, pues la predicción puede ajustarse, y es parte del modelo de Bayes, pero los qualia implican algo que no puede ajustarse.
Uno podría argumentar que no existe tal posibilidad de nueva información para re-ajustar las predicciones, y así percibir las cosas de forma diferente, en tal caso diría que nos adentramos en un terreno escabroso donde corremos el riesgo de no poder falsar la teoría del cerebro predictivo teniendo en cuenta sus propios postulados de lo que debería suceder. Si efectivamente el cerebro no puede ajustar su idea “a priori” por alguna imposibilidad perceptiva de carácter biológico ¿estaríamos hablando de un “a priori” genético inmodificable por ejemplo? ¿O la teoría es incorrecta y simplemente, hay cosas que el cerebro puede percibir y cosas que no?
En todo caso, el proceso predictivo tendría que ser parte de la filogenia de la especie más que de la ontogenia y desarrollo del organismo (aunque es verdad que durante el desarrollo se aprenden múltiples aspectos de la visión). Una cosa es que el cerebro tenga determinados patrones de procesamiento de los estímulos visuales, codificados de forma genética, y otra cosa es que el cerebro haga una codificación predictiva de forma activa, a lo Bayes, es una importante diferencia conceptual. El cerebro no predice que deba ver un rostro normal en vez de hueco, simplemente el área encargada de ver rostros es la que se activa dados los estímulos visuales, y por eso no vemos un rostro hueco. De hecho existe una área definida para ello, el área fusiforme de la cara (FFA en inglés), un área en la superficie central del lóbulo temporal que contiene células que responden a las caras, lo cual denota la importancia del procesamiento de rostros en el ser humano. Decir que este carácter filogenético es un proceso de tipo bayesiano también presenta sus problemas, puesto que el principal motor que hace que percibamos las cosas de una forma en particular es la selección natural, que ha hecho que captemos de manera más acertada o con más detalles e información las cosas que se encuentran afuera en el mundo. Aplicamos de forma errónea nuestra psicología daría a un hecho como la evolución, donde no se espera nada, no hay un a priori que luego se ajusta a la evidencia, simplemente hay características que se adaptan mejor al ambiente, y las que no desaparecen. Difícilmente podemos decir que el cerebro espera que una fuente de luz provenga de arriba (el sol), sino que las características que reaccionaron frente a este hecho se consolidaron en la evolución de la especie en particular, de modo que luego se procesan las cosas de una forma en particular, pero uno no puede ajustar la percepción para hacerla más acertada, son aspectos estructurales no modificables los que gobiernan. Si hablamos de un proceso bayesiano, el a priori debe de poder modificarse, de modo que nuestra percepción sería distinta, y quizás no existirían justamente este tipo de ilusiones ópticas.
Por lo que tal como indicaba en el anterior ensayo mencionado, debemos ser precavidos en las conclusiones que extraemos de este tipo de ilusiones ópticas. Podríamos hasta decir que lo que el cerebro hace no es predecir, sino que se confunde la predicción con la simple activación de ciertos patrones neuronales, o sea, frente a ciertas cosas se activarán ciertos patrones con más probabilidad, estos refieren a la experiencia consciente, y la activación previa de ciertos patrones hará que estos sean activados a su vez con mayor probabilidad, eso no implica un acto predictivo constante en base a lo percibido momento a momento, sino que más bien existe una percepción que condiciona la percepción, pero predecir sería algo diferente, un proceso más. Que uno esté constantemente haciendo predicciones en base a lo percibido sería ineficiente también, pues con lo otro es suficiente. A su vez hay cosas que no somos capaces de percibir, y si este fuese el caso para rostros huecos, ese hecho aislado explicaría la ilusión y no los conceptos de Bayes. Y de nuevo, no olvidemos la práctica ausencia de otras ilusiones de diferentes modalidades sensoriales, que diría serían esperables y generaría mayor soporte a la hipótesis del cerebro y la conciencia como algo predictivo.
Referencias
Adelson’s Checker-Shadow Illusion. (n.d.). Recuperado de https://www.illusionsindex.org/ir/checkershadow
Adelson, E. H. (2000). “Lightness perception and lightness illusions” in The New Cognitive Neurosciences, M. Gazzaniga (Ed), 2nd ed. pp. 339–352, MIT Press: Cambridge MA.
Felleman, D. J., & Van, D. E. (1991). Distributed hierarchical processing in the primate cerebral cortex. Cerebral cortex (New York, NY: 1991), 1(1), 1-47.
Gilchrist, A. L. (2010). “Lightness Constancy” in Goldstein, E. B. (Ed.). (2010). Encyclopedia of perception (Vol. 1). Sage.
Hills, Peter J., Holland, Andrew M., & Lewis, Michael B. (2010). Aftereffects for Face Attributes with Different Natural Variability: Children Are More Adaptable than Adolescents. Cognitive Development,25(3), 278-289.
Kandel, E. R, Schwartz, J. H. & Jessel, T. M.(2001). Manual de Neurociencia. Madrid: McGraw-Hill.
Kafaligonul, H. (2014). Vision: A Systems Neuroscience Perspective.
Kingdom, F. A. (2011). Lightness, brightness and transparency: A quarter century of new ideas, captivating demonstrations and unrelenting controversy. Vision research, 51(7), 652-673.
Ramachandran, V.S &Hirstein, W. (1997). Three Laws of Qualia: What Neurology Tells Us about the Biological Functions of Consciousness, Qualia and the Self.Journal of Consciousness Studies, 4, 429-458.
Tangen, J., Murphy, S., & Thompson, M. (2011). Flashed Face Distortion Effect: Grotesque Faces from Relative Spaces. 40(5), 628-630.
Webster, M., & Maclin, A. (1999). Figural aftereffects in the perception of faces. Psychonomic Bulletin & Review,6(4), 647-653.
Discover more from Sinapticas
Subscribe to get the latest posts sent to your email.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
One thought on “La neurociencia y las ilusiones ópticas”